Projektbeskrivelse
Et globalt perspektiv på opløst organisk stof
og verdens mindste organismer
Stiig Markager, Morten Søndergaard, Mathias Middelboe og Colin Stedmon
Abstrakt:
Stofomsætningen i havet er domineret af meget små organismer og opløste organiske stoffer (DOM). I dette projekt vil vi udnytte det globale perspektiv i Galathea 3 til en sammenhængende undersøgelse af fordeling, sammensætning og omsætning af DOM og de mikroorganismer som styrer omsætningen DOM. DOM er oceanernes største pulje af organisk kulstof og dens nettoomsætning er afgørende for den globale kulstofbalance. Vi vil bruge optiske og kemiske analyser i studiet af DOM og have fokus på, hvordan de mikrobielle samfund domineret af virus, bakterier og små alger i interaktion med fotokemiske processer bestemmer DOM puljens størrelse, sammensætning og omsætningstid.
|
Baggrund og formål Omkring 97% af alt organisk bundet kulstof i verdenshavet er i opløst form. Levende organismer, fra virus og bakterie op til hvaler, stå for under 1%, mens omkring 2% udgøres af døde partikler. Mængden af kulstof bundet i opløst organisk stof (DOM) tilsvarer mængden af kuldioxid i atmosfæren, men man ved meget lidt om de forhold, som styrer dannelse og omsætning af DOM i oceanerne, og dermed hvad der bestemmer koncentrationen. Puljens kolossale størrelse betyder, at selv små forskydninger vil påvirke det globale kulstofkredsløb. Således vil en 1% stigning i den årlige netto nedbrydning af DOM i havet betyde en større produktion af kuldioxid, end den mængde der kommer fra menneskeskabte kilder (Hedges 2002).
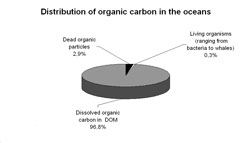
DOM i havet stammer fra nedbrydning af organisk stof på land, som ender i havet via floder, og fra organisk stof produceret af havets planter, især fytoplankton. I havet nedbrydes DOM fotokemisk og af bakterier. DOM puljens koncentration
og sammensætning varierer med afstanden fra land, dybde og
antagelig breddegrad, men man ved meget lidt om DOM puljens sammensætning, andet end at det er en kompleks blanding af en lang række stoffer, hvoraf hovedparten ligner de humusstoffer vi finder i jord. Over 80% af kulstoffet i DOM er således bundet i ukendte stoffer. Der findes kun få undersøgelser som kemisk beskriver DOM-puljen (Hansell & Carlson 2002).
Når man kigger på organismer i oceanerne opdager man, at både med hensyn til antal, vægt og aktiv biologisk overflade er det organismer under ca. 2 mm i diameter (pico- og femtoplankton), der er totalt dominerende. Det betyder ikke at store organismer er uden økologisk betydning, men det er de små alger, bakterier, virus og protozoer, der står for den største del af havets stofomsætning. Typisk vil de små algers andel af den samlede primærproduktion være mellem 30 – 80 % og med den største andel i varme regioner og næringsfattigt vand (Søndergaard 1994). Samtidig omsættes den største del af primærproduktionen omsættes af bakterier via DOM (Williams 2000), og marine virus udgør en væsentlig tabsfaktor for bakterieplanktonet (Fuhrman & Noble 1995). Der er derfor en tæt kobling mellem dynamikken af DOM og de mikrobielle organismer og processer.
Det fremgår af den nyeste internationale forskning (Kirchman 2000, Hansell & Carlson 2002), at der findes geografiske og temporære forskelle i ovennævnte relationer, men systematikken i disse sammenhænge over klimatiske zoner og i forskellige typer af havvand er dårligt undersøgt. Galathea 3 ekspeditionen giver en enestående mulighed for et globalt perspektiv og for at opnå ny indsigt i betydningen af DOM samt de mikrobielle processer og fysisk-kemiske faktorer, der styrer dets produktion og nedbrydning og driver store dele af klodens biogeokemiske kredsløb. Det særlige er at det er det samme skib, og dermed samme instrumenter, protokoller og ansvarlige forskere, som udfører målingerne hele jorden rundt. Dermed elimineres de usikkerheder som opstår ved sammenligning at resultater fra forskellige undersøgelser, og der kan skabes et datasæt som os bekendt ikke er opnået før.
Relation til tidligere forskning og perspektiver
Karakterisering af DOM puljen DOM puljen er meget svær at karakterisere kemisk, bortset fra ”bulk” indholdet af kulstof (DOC), kvælstof (DON) og fosfor (DOP). Derfor har vi i ansøgergruppen i de senere år arbejdet med en optisk metode til karakterisering af DOM, og med DOM-puljens funktionelle egenskaber i form af følsomhed over for fotokemisk og bakteriel nedbrydning.
|
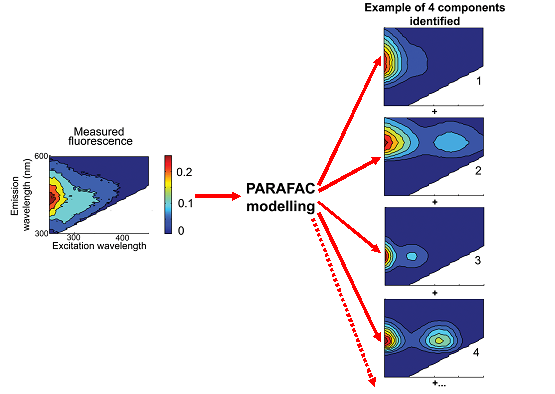
Fig. 2. Figuren viser fluorescensen fra opløst organisk stof i en vandprøve. Prøven belyses med forskellige bølgelængder (vandret akse) og fluorescensens intensitet registreres (farvekonturer) over hele spekteret (lodret akse). Ved hjælp af en statistisk model (PARAFAC) opdeles signalet i komponenter som svarer til stofgrupper i DOM-puljen.
|
DOM puljens sammensætning kan undersøges med en optisk metode baseret på fluorescens egenskaber. Metoden kan på få minutter karakterisere DOM puljen i en prøve (Stedmon et al. 2003). Metoden giver ikke en kemisk karakteristik af DOM, men beskriver den relative fordeling af en række komponenter i DOM puljen. Metoden har indtil nu været anvendt i flere studier af dynamikken i DOM puljen i fjorde, åbent hav og søer på Antarktis (Fulton et al 2004; Stedmon og Markager 2005a,b; Stedmon et al. indsendt). Målinger af 3-dimensionelle fluorescensspektre har tidligere været anvendt andre steder i verden, fx Coble (1996), men problemet har hidtil været at fortolke og kvantificere resultaterne. Dette problem er nu løst ved anvendelse af den multivariate statistiske metode PARAFAC. Et globalt datasæt fra Galathea 3 vil gøre det muligt at udnytte metodens fulde potentiale inden for et område med stor international bevågenhed.
Den optiske metode åbner mulighed for at undersøge både dynamikken i produktion og nedbrydning af DOM. I nærheden af kyster og floder er DOM-puljen variabel og afhængig af det som kommer lokalt fra land og den lokale produktion i havet. I det åbne hav er de processer som styrer produktion og nedbrydning mere ensartede, dog med mulighed for en variation med breddegraden relateret til temperatur, indstråling og næringsforhold. Man kan derfor opstille den hypotese, at DOM-puljen vil konvergere, fra en meget variabel sammensætning kystnært, hen mod en standard sammensætning i de åbne oceaner – at der findes en fælles global pulje af oceanisk humus eller ’havets jord’ om man vil. Galathea 3 togtet giver en enestående mulighed for at indsamler et globalt datasæt, som gør det muligt at teste denne hypotese.
DOM – nedbrydning og bakterier DOMs variation i koncentration som funktion af biologisk aktivitet og følsomhed over for fotokemisk og bakteriel nedbrydning er et andet nyt og stort forskningsområde (Carlson 2002). Indtil for ca. 20 år siden troede man, at hovedparten af DOM-puljen i havet var refraktiv, dvs. stort set ikke nedbrydelig. I dag ved vi, at det ikke er korrekt, og at nedbrydningen sker i et kompliceret samspil mellem fotokemiske processer og bakterier (bl.a. Søndergaard & Middelboe 1995). Tidsskalaen for nedbrydning af DOM bestemmer hvor lang tid den CO2, som bindes i planteplanktonets produktion, er om at vende tilbage til atmosfæren. Nedbrydningen af DOM er også vigtig for forståelsen af kvælstof- og fosforkredsløbet i havet, idet langt hovedparten af alt ikke atomart kvælstof og de største mængder fosfor i havet er bundet i DOM. Vi foreslår derfor, at man under togtet udfører en række eksperimenter, hvor nedbrydningen af DOM følges lang tid (måneder), samtidig med at vi måler ændringer i DOM-puljens sammensætning, iltforbrug, frigivelse af næringssalte og bakteriel aktivitet og følsomheden over for ændringer i temperature. Forsøgene udføres om bord under kontrollerede betingelser.
Kun bakterier kan udnytte DOM til vækst. I forskellige oceaniske regimer synes den bakterielle aktivitet og dermed bionedbrydningen af DOM at være kontrolleret enten af næringssalte (Carlson et al. 2002), biotilgængeligheden af DOM (Kirchman 1990) eller af biotiske faktorer (Søndergaard et al. 2000). Nedbrydningsforsøgene over længere tid vil derfor blive suppleret med korttidsforsøg, hvor det undersøges, hvad der aktuelt begrænser den bakterielle aktivitet. Forsøgene udføres i en forsøgsmatrix med forskellige temperaturer, hvor der tilsættes uorganiske næringssalte og organiske substrater og den bakterielle produktion og tilvækst måles over tid.
Virus Som omtalt spiller virus en afgørende rolle i reguleringen af den bakterielle aktivitet i havet og dermed for omsætningen af DOM. Viral lysis af heterotrofe og autotrofe mikroorganismer frigiver letomsætteligt, næringsrigt cellemateriale, som dermed igen bliver gjort tilgængeligt for andre, ikke-inficerede bakterier (Furhman 1999, Middelboe et al. 2003). Virale lysater udgør således potentielt en vigtig fødekilde for bakterier. En del af det organiske materiale i havet cirkulerer derfor mellem bakterier og puljen af DOM som følge af viral aktivitet. Denne ”virale løkke” stimulerer den mikrobielle respiration i vandsøjlen og reducerer transporten af kulstof til højere trofiske niveauer i den marine fødekæde (Fuhrman 1999). Da omkring 75 % af det frigivne organiske materiale ved cellelysis udnyttes indenfor få timer (Middelboe et al. 2002) er det givet, at disse lysater kan udgøre et kvantitativt vigtigt bidrag til det bakterielle kulstof- og næringsstofbehov i havet. I nogle tilfælde har virale lysater endda vist sig at udgøre hoved-fødekilden for bakterierne (Wilhelm & Suttle 1999). Endnu ved vi ikke meget om den kvantitative betydning af den virus-inducerede regenerering af organisk materiale, eller hvad der regulerer den relative betydning af denne proces i forhold til andre bidrag af omsætteligt organisk materiale i et givent havområde. Det er derfor formålet at undersøge betydning af virale processer på omsætning af kulstof og næringssalte i forskellige havområder, der er karakteriseret ved forskelle i bl.a. produktivitet, temperatur og sammensætning af de autotrofe samfund. På baggrund af sådanne undersøgelser vil vi kunne vurdere virus’ rolle i omsætningen af kulstof og næringssalte på såvel regional som på global skala. Dette er en afgørende, og manglende, brik i en kvantitativ forståelse og beskrivelse af det mikrobielle stofkredsløb i havet.
Ud over at bestemme antal og fordelingen af virus i vandsøjlen i relation til bakterieantal og aktivitet vil der på de udvalgte lokaliteter blive gennemført eksperimenter ombord på skibet. Eksperimenterne skal belyse betydningen af virus for dannelse og omsætning af DOM. Denne type eksperimenter har alle været afprøvet, testet og udnyttet i andre studier (bl.a. Søndergaard et al. 2000, Middelboe & Lyck 2002, Middelboe et al. 2002, 2003).
Picoalger Først i 1979 blev det erkendt, at små cyanobakterier (1-2 mm) af slægten Synechococcus kan være de dominerende primærproducenter i oceanerne (Waterbury et al. 1979). I de efterfølgende år blev det klarlagt, at små fotosyntetiserende organismer findes i næsten alle undersøgte marine og ferske vande, men er mest dominerende under varmere himmelstrøg og i næringsfattigt vand (Søndergaard et al. 1991, Søndergaard 1994). Det er også velkendt at picoalger fordeler sig på en forudsigelig måde i vandsøjlen med gruppen prochlorophyter i bunden af den fotiske zone, men tilsyneladende kun under varmere himmelstrøg (Chrisholm et al. 1988). Udforskningen af picoalgernes globale fordeling er ikke forbi (Sherr & Sherr 2000), og der er kun gennemført ganske få komparative studier, hvor deres globale fordeling såvel horisontalt som rumligt sammenkædes med andre organismer i det mikrobielle fødenet og til DOM. Et sådan studie vil betræde ”nyt vand” og kunne yde et væsentligt forskningsmæssigt bidrag til forståelsen af de mindste organismers betydning og sammensætning over store geografiske og klimatiske gradienter, over næringsgradienter (upwelling regioner), under næringsfattige forhold i de tropiske oceaner samt under mere ekstreme betingelser ved Antarktis.
Der er foretaget mange målinger af DOM og mikrobiel aktivitet i oceanerne, men sjældent med det globale perspektiv Galathea 3 giver mulighed for, og os bekendt aldrig med samtidige undersøgelser af DOM puljens størrelse (DOC, DON og DOP koncentrationer), sammensætning og omsætning og kvantificering af virus. Vi vil udnytte alle muligheder, hvor der tages overfladevand og dybdeprofiler til at få kvantificeret DOM, picoalger, bakterier og virus. Samtidig udtages prøver til måling af bakteriel produktion (leucin og thymidin metoderne). Måling af den bakterielle produktion vil give et overordnet udtryk for den mikrobielle omsætning af DOM puljen. Hvis det er teknisk muligt på skibet kan fluorometeret kobles direkte til skibets stævnrørsindtag, således at DOM måles kontinuert mens skibet sejler.
Placeringen af et flowcytometer om bord vil give mulighed for en næsten ”on line” bestemmelse og kvantificering af flere typer picoalger (cyanobakterier, eukaryoter, prochlorophyter) samt tælling af de mest dominerende større planktonalger. Efter farvning af vandprøver med fluorescerende DNA og RNA farvestoffer (SYBR green 1 og gold) kvantificeres bakterier og virus ligeledes ved tælling i flowcytometer, hvilket betyder at mange forsøgsresultater bliver analyseret om bord. For bakterier er det muligt at skelne mellem celler med højt og lavt indhold af DNA, dvs at få kvantificeret antallet af hurtigt voksende bakterier (Søndergaard 2000) og identificere op til 4 grupper virus, hvor algevirus kan adskilles fra andre virus.
Det er vores vurdering, at vi med det planlagte prøvetagningsprogram og eksperimenterne udført om bord kan opnå videnskabelige landvindinger med et globalt perspektiv; noget der mangler i mange af de ”cruises”, der foretages rundt om på verdenshavene.
Opsummering af formål:
- at gennemføre måleserier, der viser den globale fordeling DOM i havet
- at karakteriserer DOM-puljen med nye optiske metoder og teste en hypotese om at den konvergerer hen mod en fælles oceanisk pulje
- at koble de kemiske og optiske målinger af DOM med sammensætningen af de mikrobielle samfund (bakterier, virus og picoalger) og deres aktivitet
- at gennemføre eksperimenter i udvalgte havområder (klimatiske gradienter og næringsgradienter), der vil skaffe ny viden om, hvor hurtigt DOM omsættes, hvilke forhold der begrænser bakteriers aktivitet og kvantificere betydningen af virus for havets DOM omsætning
- at anvende den opnåede viden til en analyse af klimaforandringers effekt på omsætningen af DOM
Internationalt samarbejde De tre deltagende institutioner har en lang tradition for internationalt samarbejde i form af deltagelse i projekter i nordisk og EU regi og ved udveksling af medarbejdere, Post Doc. og studerende. Konkret kan nævnes at DMU samarbejde med fire universiteter i USA og Canada og firmaet WetLab Inc. om udviklingen og anvendelse af metoden med fluorescens karakteristik. DMU arrangerede også en workshop om emnet i marts 2005 med deltagelse af 10 institutioner fra 5 lande, og har pt. udveksling af medarbejdere med to universiteter i Canada inden for dette område. Desuden samarbejdes med Uppsala Universitet og Åbo Akademi, Finland om nedbrydning af DOM i Østersøen. Københavns Universitet er pt. i gang med planlægningen af en udveksling af medarbejdere med Flinders University, Australien. Mathias Middelboe er desuden medlem af en arbejdsgruppen om virus under Scientific committee for Oceanographic Research. Vi vil undersøge mulighederne for at inviterer udenlandske samarbejdspartnere med på togtet når vi ved om projektet bevilges og aktuelle tidspunkter.
Kvalifikationer og tidligere arbejde Forskergruppe har gennem de seneste år udviklet nye optiske metoder og en avanceret statistisk model til karakterisering af DOM puljen (Stedmon et al. 2003), præget den internationale udforskning af virus funktionelle rolle for havets stofomsætning (Middelboe et al. 2002, 2003) og stået i spidsen for store internationale undersøgelser af DOM produktion og nedbrydning i flere typer akvatiske økosystemer (Søndergaard & Thomas 2004, Kragh & Søndergaard 2004). Vi mener således at vi tilsammen har den nødvendige faglige ekspertise og erfaring til at gennemføre det foreslåede projekt.
Arbejdsplan Et af de unikke aspekter ved dette projekt er, at det dækker globalt, og planen er derfor at det gennemføres kontinuert under hele togtet. Analyser som udføres automatisk som fx. DOM fluorescens, flowcytometri og DOC koncentration vil blive udført på så vidt muligt alle vandprøver. Dette kan udføres ved deltagelse af en person fra gruppen. De mere arbejdskrævende inkubationer vil blive udført på så mange prøver som muligt og kræver tilstedeværelsen af 2 personer. Hvis det ikke er muligt at finde plads til 2 personer fra projektgruppen på hele togtet, er de fagligt mest interessante områder for nedbrydningsforsøg Nordatlanten, Middelhavet, de indonesiske havområder, Salomonøerne, havet omkring Antarktis og ud for Peru og det Caribiske Hav.
Formidling Det er vores ambition at formidle betydningen af DOM og havets mikroorganismer til et bredt publikum. Under togtet vil det ske med hjælp fra de medfølgende professionelle formidlere. Desuden forestiller vi os at resultaterne fra både flowcytometer og fluorescensmålinger formidles undervejs til fx skoleklasser og via togtets hjemmeside. Begge instrumenter giver resultater straks, og de kan, med den rette tilpasning og forklaring, formidles som ’farvestrålende’ figurer (noget hen ad Fig. 2, men forenklet). Resultater fra fluorescensmålinger kan fx vise at man nærmer sig land pga. af en stigning i fluorescensen DOM, længe før det i øvrigt kan registreres med andre metoder eller ses med det blotte øje. Det er også muligt at lave beregningsprogrammer som er så simple, at fx skoleklasser (gymnasiet) kan bruge dem til at behandle de data der indsamles undervejs, således at de kan følge med i hvilke typer af DOM skibet møder og vandets indhold af alger, bakterie og virus. Her forudsættes at datakommunikationen tillader at data hjemsendes elektronisk. Vi tror, at vi på den måde kan formidle essensen af den naturvidenskabelige arbejdsmetode til et bredt publikum ved at kombinere den seneste teknik, Galatheas global perspektiv, et skær af eventyr og en væsentlig miljømæssig problemstilling. De videnskabelige resultater vil naturligvis også blive publiceret internationalt og i danske populærvidenskabelige tidsskrifter. Det vil helt overvejende ske efter togtet.
Referencer
Carlson, C. A. 2002. Production and removal processes. In: Hansell, DA & CA Carlson. 2002. Biogeochemistry of marine dissolved
organic matter. Academic Press. Pp. 91-151.
Carlson, C. A. et al. 2002. Effect of nutrient amendments on bacterioplankton production, communitystructure, and DOC
utilization in the northwestern Sargasso Sea. Aquat. Microb. Ecol. 30: 19-36.
Chrisholm, S.W. et al. 1988. A novel free-living prochlorophyte abundant in the oceanic euphotic zone. Nature 334:340-343.
Coble, P.G. 1996. Characterisation of marine and terrestrial DOM in seawater using excitation-emission matrix spectroscopy. Marine
Chemistry, 51:325?346.
Fuhrman, J. A. 1999. Marine viruses and their biogeochemical and ecological effects. Nature 99:541-548.
Fuhrman, J.A. & R.T. Noble. 1995. Viruses and protists cause similar bacterial mortality in coastal seawater. Limnology and
Oceanography 40: 1236-1242.
Fulton, J.R. et al. (2004) Changes in fulvic acid redox state through the oxycline of a permanently ice-covered Antarctic lake. Aquat.
Sci. 66, 27-46.
Hansell, DA & CA Carlson. 2002. Biogeochemistry of marine dissolved organic matter. Academic Press.
Hedges, J.I. 2002. Why dissolved organics matter? In Hansell, D.A. and Carlson, C.A., (Eds.) Biogeochemistry of Marine Dissolved
Organic Matter. Academic Press.
Kirchman, D. L. 1990. Limitation of bacterial growth by dissolved organic matter in the subarctic Pacific. Mar. Ecol. Prog. Ser. 62:
47-54.
Kirchman, D. L. (ed). 2000. Microbial ecology of the oceans. Wiley-Liss, New York.
Kragh, T. & Søndergaard, M. 2004. Production and bioavailability of dissolved organic carbon: effects of mesozooplankton.
Aquat. Microb. Ecol. 36: 61-72
Middelboe, M., and P. G. Lyck. 2002. Regeneration of dissolved organic matter by viral lysis in marine microbial communities.
Aquat. Microb. Ecol. 27:187-194.
Middelboe, M., T. G. Nielsen, and P. K. Bjørnsen. 2002. Viral and bacterial production in the North Water polynya: In situ
measurements, batch culture experiments, and characterization and distribution of a virus-host system.
Deep Sea Research II 49:5063-5079.
Middelboe, M., L. Riemann, G. L. Steward, V. Hansen, and O. Nybroe. 2003. Virus-induced transfer of organic carbon between
marine bacteria in a model community. Aquat. Microb. Ecol. 33:1-10.
Sherr, E. & B. Sherr. 2000. Marine microbes: An overview. In: Kirchman, D. L. (ed). 2000. Microbial ecology of the oceans.
Wiley, New York, pp. 13-46.
Stedmon C.A. and S. Markager (2005)a. Resolving the compositional changes in dissolved organic matter (DOM) in a temperate
estuary and its catchment, using spectrofluorometry and PARAFAC analysis. Limnol. Oceanogr. 50: 686-697.
Stedmon C.A. and S. Markager.(2005)b Tracing the production and degradation of autochthonous fractions of dissolved organic
matter using optical analysis. Limnology & Oceanography 50: 1415-1426.
Stedmon, C., Markager, S and R. Bro. (2003): Tracing dissolved organic matter in aquatic environments using a new approach to
fluorescence spectroscopy. Mar. Chem. 82, 239-254.
Søndergaard, M. et al. 1991. Picoalgae in Danish coastal waters during summer stratification. Mar. Ecol. Prog. Ser. 79:139-149.
Søndergaard, M. 2000. Bacterioplankton: How many and how active? Verh. Int. Ver. Theor. Angew. Limnol. 27: 859-865.
Søndergaard, M. 1994. Picoalger, de mindste primærproducenter. Vand & Jord 5: 199-203.
Søndergaard, M. & M. Middelboe. 1995. A cross-system analysis of labile dissolved organic carbon. Mar. Ecol. Prog.
Ser. 118:283-294.
Søndergaard, M. & D. Thomas (eds). 2004. Dissolved organic matter (DOM) in aquatic ecosystems. (www.domaine.ku.dk).
ISBN 87-89143-25-6.
Søndergaard, M. P. J.leB Williams et al. 2000. Net accumulation and flux of dissolved organic carbon and dissolved organic nitrogen
in marine plankton communities, Limnol. Oceanogr. 45: 1097-1111.
Waterbury, J.B. et al. 1979. Widespread occurrence of a unicellular, marine planktonic cyanobacterium. Nature 277:293-294
Wilhelm, S. W., & C. A. Suttle. 1999. Viruses and nutrient cycles in the sea. Bioscience 49:781-788.
Williams, P. J. leB. (2000) Heterotrophic bacteria and the dynamics of dissolved organic material. In: Kirchman, D. L. (ed). 2000.
Microbial ecology of the oceans. pp. 153-200.
|