Områdets oceanografi
Physical Oceanography of the Western Australian Shelf.
Leeuwin Current.
The mid latitude oceans lying off the west coast of continents are extremely productive. The classic example is off the coast of Peru and Chile; other examples include the waters off Namibia, California and Portugal; and so too Western Australia. In most of these cases, the primary driver of this productivity is wind driven upwelling. Eastward trade-winds at these latitudes drive warm waters off shore bring deep, nutrient rich waters to the surface so fueling the growth of phytoplankton. This, however, is not the mechanism driving production on the west Australian shelf. Western Australia is unique in this sense, in that it is the only example in the world where there is a persistent pole-ward eastern boundary current, carrying warm tropical waters towards the southern ocean. This is the Leeuwin Current. It is driven not by winds, but by density differences: warm and relatively fresh (i.e. light) water in the north and cold, salty (i.e. heavier) water in the south. These warm waters flow southward over the cooler saltier deep counter current flowing northward. This flow more than overwhelms the ability of the trade winds to bring deep waters up towards the surface.
Internal Waves:
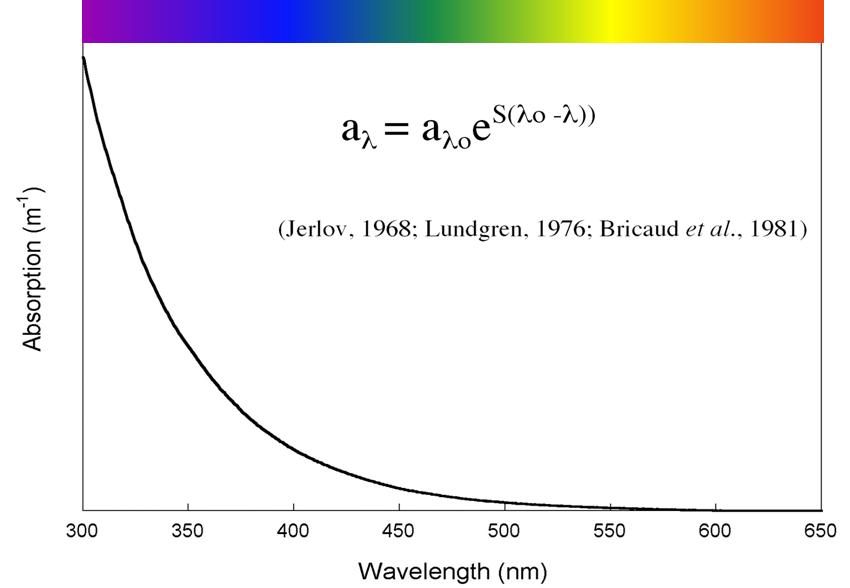
So where does the energy come from to stimulate the observed primary production? One likely candidate is mixing mediated by internal waves. It is known that up to 30% of the worlds tidal energy goes into internal waves; transporting this energy thousands of kilometers from where it was “removed” from the tide, to where it finally dissipates in turbulent mixing. The attached figure shows regions where tides loose energy; in those areas away from coasts (Madagascar Plateau, Kerguelen Rise, South West Indian Ridge, and the North West Australian shelf) this energy is lost to internal waves. Similar transfers of energy to internal waves occur in the same locations from wind driven currents. Internal waves are generated wherever there is a flow over a bathymetric feature; (a sea mount, or ridge for instance), and where there is a change in water density with depth (i.e. stratified). Provided the oceans remains stratified, these internal waves can travel vast distances, bouncing from surface to bottom, with very little loss of energy. That is, at least, until very special conditions are met. Depending on how the water column is stratified and the frequency at which the internal waves are generated, there is critical bottom slope where all of the energy of the wave becomes focused into a very small volume. This causes the wave to break, generating turbulence, and mixing deep nutrient rich waters into the surface of the ocean. This is thought to happen along the coast of Western Australia. There are indeed internal waves observed there from satellites (see opposite) in this area. The question we seek to address is, are these persistent and important in biological processes that play out on the Western Australian shelf.
|
|
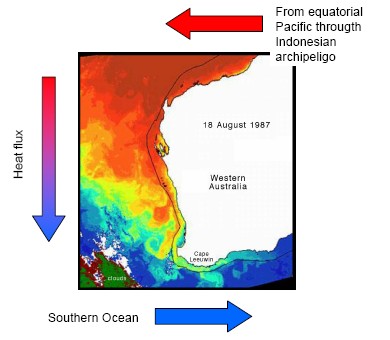
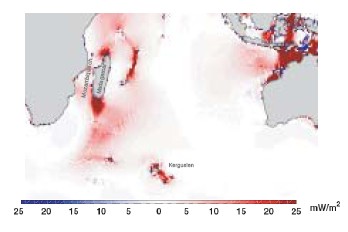
Red areas show where internal tides are generated.
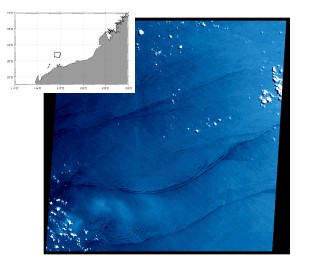
Satellite image of the surface expression of internal waves off the coast of Broome, Western Australia.
|